

The Push-me, pull-you of plant water: circulation without a heart
The Push-me, pull-you of plant water: circulation without a heart
Plant and ecosystem resilience is built from diversity – diversity within individuals, diversity between individuals and across species.


A couple years ago I planted a Redbud tree in my front yard. Every spring it flowers well before the leaves come out. It always kind of baffled me. Recently, I finally understood this ability to flower before photosynthesis started up again must have been stored in the tree all winter long. Come to find out surplus sap (non-structural, dissolved, photosynthesized carbon) and water collect in the tree as it prepares for winter and beyond. In spring, when the temperatures rise, the sap and water squeeze out the tree to form flowers.
It is not just flowers the tree prepares for by storing non-structural carbon. Trees store enough excess carbon to re-foliate their entire canopy without photosynthesis. This storage system helps to protect the tree against down years (Muhr, et.al., 2016).
In case of hard times, trees (and other plants) also store water in their rooting zones. This is storage built by the tree but outside the tree. The importance of this ‘rooting zone storage’ grows as the mismatch in timing between the rainy season and growing season grows. Like in a Mediterranean climate, where winter brings rain and the growing season is dry. In these environments and in temperate and tropical climates experiencing droughts, the only water potentially available for photosynthesis has to be already stored in the ground.
Available water means “reachable” by the plant – reachable by proximity but also “absorbable”. By design, the routes by which water gets to the plant are many. This includes the many, distributed fine root absorption areas to suck up moisture, but also the connections between tree individuals and others, like mycorrhiza fungi, that can pass on water to the tree. Water can also be absorbed on the outside of the tree, with the help of other fungi, lichen and mosses. For coastal redwoods water in the form of fog can be absorbed through the bark of twigs and through the leaves.
The internal architecture of plants, the rooting zone and the atmosphere have evolved together to move and store water and are connected through the plant’s vascular system. This vascular system is like ours but made of living and non-living cells - tracheids and vessels. The inside of plants, like soils, is a complex porous media built by plants with parallel, redundant points of entry, bi-directional flows and dynamic storage.
Research has shown that the water and sap in a plant can have a wide variety of ages (for example, see: Muhr above, Nehemy, et.al., 2022 and Bowers and Williams, 2022). The living and dead cells that compose the plant water highways can become partially or completely air-filled and fail. Some flow pathways get cut off and some cut off paths get re-hydrated and restored. The plant itself is a living, dynamic flow network just like hillslope flow networks are.
There are water conducting cells in the phloem and xylem. The phloem is typically thought of as conducting dissolved, photosynthesized carbon down from the leaves and the xylem as bringing water up from the roots. But, of course, it is not that simple. The phloem and xylem are hydraulically connected radially by parenchyma rays. These connections, like all the plant and rooting zone flow routes accommodates bi-directional flows.

The phloem does conduct photosynthetic products down from the leaves, but it is also an elastic storage. Stems, just like roots, undergo small but significant changes in diameter diurnally as the tree breathes. During photosynthesis water is pulled/supplied by the phloem to the xylem through the rays and the stem diameter contracts. At night, the dried-out phloem cells are replenished by xylem and root water, and the stem diameter expands.
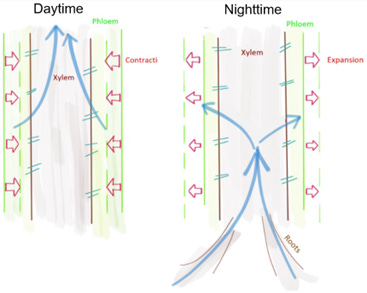
This is a form of water redistribution driven by a water potential gradient that does not include evaporation. This gradient has less energy than evaporation but enough vapor, hydrostatic and osmotic pressure to move water from wetter areas, even against gravity, to drier areas. This plant-associated water - hydraulic redistribution was first noticed in phreatophytes (groundwater-loving) woody plants (Prieto, et.al. 2011). These plants can have deep roots, frequently tap roots that reach deep into the ground.
At night these woody plants draw up groundwater or other wet areas via deeper roots releasing it back through shallower roots into drier soil reserves. Water is passively conducted through this root system. It could be a vapor pull or osmotic push or a combination. Typically, this lifted water is only a small proportion of the overall water that a plant transpires annually but can sometimes be a significant fraction (Prieto, et.al., 2011). This was first called hydraulic uplift. But now it is known that water can move all around the plant based on non-evaporation driven water potential gradients.
The old theory of transpiration is that evaporation pulls on a continuous column of water in the plant from top to bottom. But I believe more in the ‘water gate’ theory (Zimmerman, et.al., 2004). This theory does not rely on a continuous column of water but on a continuous chain of events, that lifts water up a tree, like locks lift ships up a waterway. I don’t think the water gate theory negates the potential occurrence of a hard pull on a long column of water. Maybe that happens frequently. But clearly plants do not accomplish water-related tasks without some redundancy and parallel-pathing for getting things done.
As the blossoms fade in my Redbud, the leaves come out. When the weather turns and the sun bears down during the growing season, the atmosphere pulls on water everywhere: the ocean, fresh water, from the ground and from animals and plants. The force of evaporation, coupled with growth exerts a water suction in the plant all the way down to the roots, all day long. That suction pulls water from out of the plant and water up from the ground through the plant.
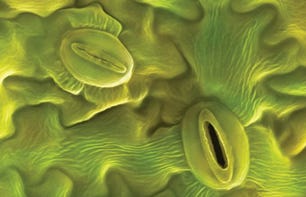
Transpiration is evaporation at the stomata. Stomata are the openings on the underside of leaves where plants inhale and exhale. Plant breaths take in air, extract the carbon and give back water and oxygen. Stomata guard against (as much as they can) plant injury from that relentless evaporative suction. They are the stopgap against over-investing in growth at the expense of water. When they close the exchange of air and water and the growing mostly cease.
Getting water up from the roots, through the shoot and the branches and out through the leaves is fraught with possible failure and loss of life, one cell at a time. Plant physiologists, among others, study the hydraulic weaknesses of plant hydraulic architecture like engineers study the failures of water movement in pipes and pumps. It is the same mechanism that cause failures in both: when air gets into what was a continuous flow of water. It is like a vacuum, only this time when the failure occurs air “pops” out of the water, pitting pump impellers and pushing the water out of a cell. When that happens, the cell can no longer conduct water. If that condition persists the cell will die.
I used to think transpiration was just a wasteful outcome of photosynthesis. Now I think this is another kind of “bypass water” headed back to that big water storage in the sky. This is water plants can afford, most of the time, to return, to pass on.
Plants and their rooting zones have had to learn some hard lessons evolving with changing climates. The drier the climate the more idiosyncratic the plant adaptations and the more intense the preparations for future droughts. Two prime examples of this come from studies in in the Red Butte Canyon Research Area in the Wasatch Mountains immediately east of Salt Lake City, Utah and from the H.J. Andrews Experimental Forest from the Western Cascade Mountains of Oregon.
In Utah, Dawson and Ehleringer (1991) found that the streamside plants – scrub oak and maple trees were getting their water mainly from deeper groundwater and not from the shallow stream they grew alongside. In the Cascade Mountains Brooks, Barnard, Coulombe and McDonnell (2010) compared the concentrations of isotropic hydrogen (deuterium) water in a Mediterranean climate. They found that the isotropic concentration in the water falling from the sky and the streams were similar, but the soil water beneath ~30-year-old Douglas Fir was more concentrated because it was feeding transpiration. The water was being held in finer pores and was regulated tightly within the rooting zone.
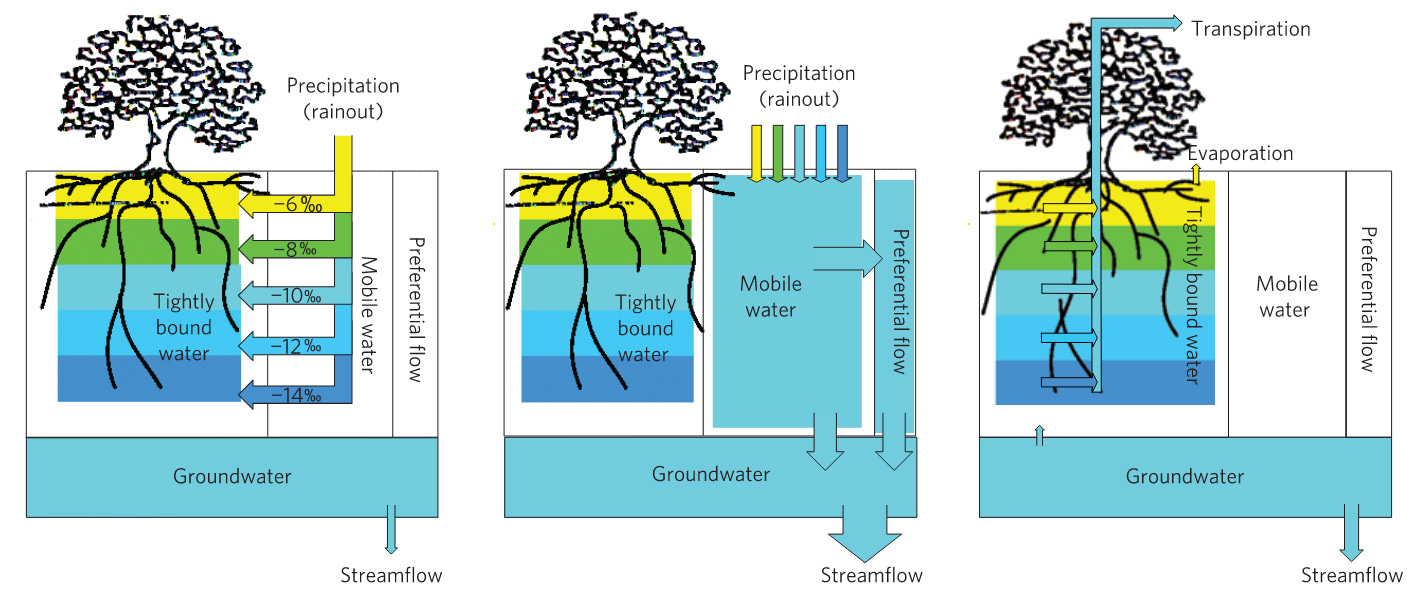
Up to the point of Brooks’ study, hydrologists tended to think that water fell from the sky, infiltrated into the ground and as it did, displaced the water that was already there. Brooks and others (for example, see: Lee et.al., 2021; Nehemy, et.al., 2020 and Liu, et.al., 2020). have consistently shown that a portion of rainfall, particularly in drier climates, is conserved in the rooting zone. It is held on to, and any rain that falls after this tightly-bound, water “storage” is filled, bypasses the storage and feeds groundwater and streams directly. This is another kind of plant-mediated, bypass flow.
Why would plants use deeper water than the water in a shallow and much easier to access stream? And why would plants choose to “pull” water from harder-to-pull-from pores than just grabbing the next rain that soaks in at the surface?
I’d say: because they have already done the prep work for hard times ahead. They have prepared for extremes and just because the weather isn’t dry at the time or the stream isn’t empty like it sometimes gets, doesn’t mean these plants are foregoing their long-evolved strategy for a transient bump. They have learned the hard way and been let down by less intensive strategies before.
The big takeaway here is that hydraulic diversity – diversity of hydraulic flow routes and storages in the plant, the diversity of ways plants acquire and store water outside their bodies, including structuring the rooting zone, along with the return of water to the atmosphere, creates the redundancy, fallbacks and fail safes that help build climate resilient individuals and ecosystems.
Now if we can also remember that the birthing process is hydraulic....thank you for the lovely read.
Fascinating. I learned a lot. Thank you